国立環境研の深谷さんと共同で進めていた論文が、Nature Communications誌に出ました。4年以上かけた、深谷さんの大作です。

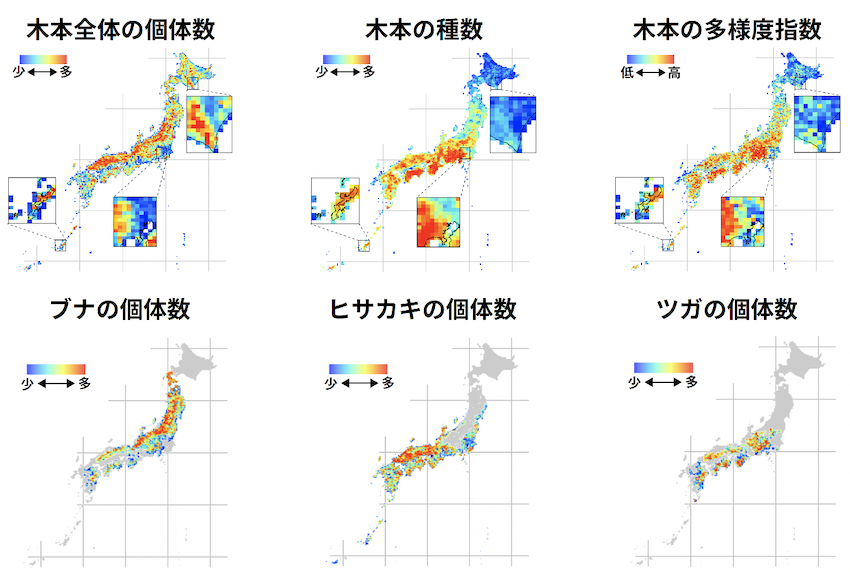
深谷さんの論文の新規的な点は、日本に分布している樹木の絶対個体数を樹種ごと(1,200種以上)、地域(約10 km平方のグリッド)ごとに推定したことです。元になったデータは、膨大な植生調査データや植物標本や植物分布の記録などです。植物の分布に関わるあらゆる情報(生物多様性ビッグデータ)です。
樹木の数を広域的に推定した先行研究(Crowther et al. 2015. Nature 525: 201–205)はあったのですが、種別の個体数を全土にわたって明らかにした研究は、深谷さんの論文が世界で初めてです。
樹木の数を広域的に推定した先行研究(Crowther et al. 2015. Nature 525: 201–205)はあったのですが、種別の個体数を全土にわたって明らかにした研究は、深谷さんの論文が世界で初めてです。
それで、「樹木の個体数が把握できたら何が凄いのか?」ということですが、これがまた面白い話になるのです。
マクロスケールの種個体数情報を元にして、今まで観測できなかったマクロ進化のパラメータ(種分化率や種の寿命など、生物多様性の起源と維持に関わる根源的なプロセス)を明らかにできたことです。
理論的には、地球上の全生物の種を捕捉して各種の絶対個体数を数え上げれば、マクロ進化に関わるパラメータを明らかにできるのですが、実際には観測不可能なので、マクロ進化パラメータを実データから推定するのは困難でした。
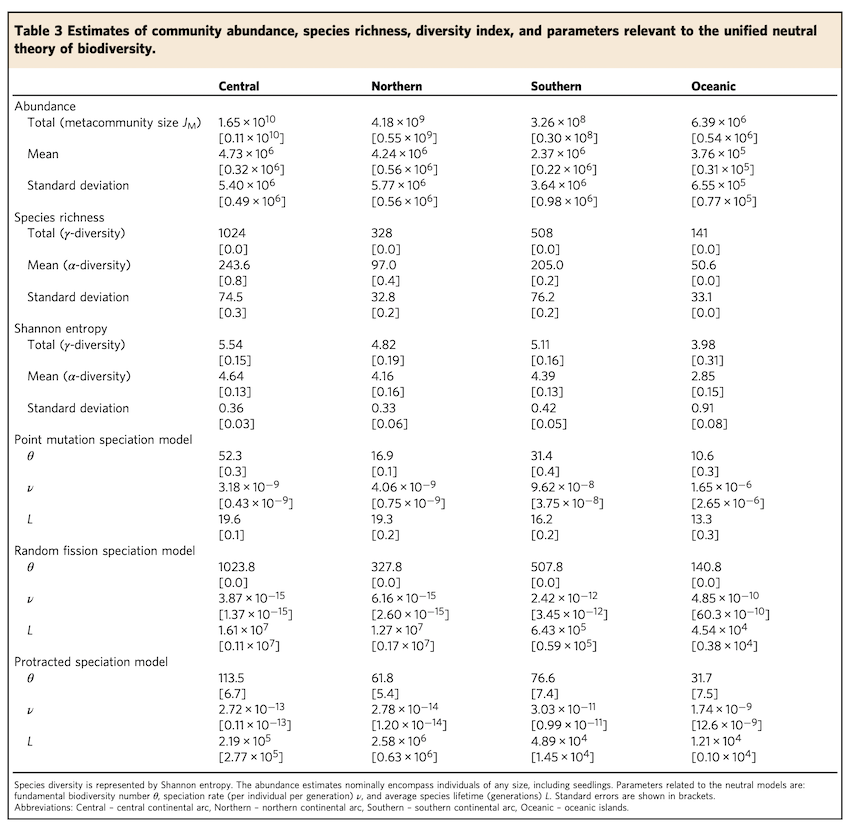
深谷さんの論文では、日本全土の自然林の樹木の絶対個体数が約210億本であることを明らかにして、約1200種の樹木種について個体数まで把握できたので、この種個体数データに中立理論を適用することで、地域ごとの樹木種の種分化率や種寿命(種が分化して生まれてから絶滅するまでの平均時間)が定量できました。
以下の表、protoracted speciation model(マクロ進化の延長種種分化モデル)をみてもらえるとわかりますが、日本中央(Central)、北海道(Northern)、琉球諸島(Southern)、小笠原諸島(Oceanic)ごとに、種分化率(ν)や種の平均的な世代数(L)が推定値が示されています。
樹木種の平均的な世代年数を30年くらいとすると、Lに30年をかけた値が、種の平均的な期待寿命になります。琉球諸島や小笠原諸島では、種の期待寿命も比較的短くなっており、歴史的に種の入れ替わり(種分化と絶滅)が盛んに生じたことがわかります。
マクロスケールの種個体数情報を元にして、今まで観測できなかったマクロ進化のパラメータ(種分化率や種の寿命など、生物多様性の起源と維持に関わる根源的なプロセス)を明らかにできたことです。
理論的には、地球上の全生物の種を捕捉して各種の絶対個体数を数え上げれば、マクロ進化に関わるパラメータを明らかにできるのですが、実際には観測不可能なので、マクロ進化パラメータを実データから推定するのは困難でした。
深谷さんの論文では、日本全土の自然林の樹木の絶対個体数が約210億本であることを明らかにして、約1200種の樹木種について個体数まで把握できたので、この種個体数データに中立理論を適用することで、地域ごとの樹木種の種分化率や種寿命(種が分化して生まれてから絶滅するまでの平均時間)が定量できました。
以下の表、protoracted speciation model(マクロ進化の延長種種分化モデル)をみてもらえるとわかりますが、日本中央(Central)、北海道(Northern)、琉球諸島(Southern)、小笠原諸島(Oceanic)ごとに、種分化率(ν)や種の平均的な世代数(L)が推定値が示されています。
樹木種の平均的な世代年数を30年くらいとすると、Lに30年をかけた値が、種の平均的な期待寿命になります。琉球諸島や小笠原諸島では、種の期待寿命も比較的短くなっており、歴史的に種の入れ替わり(種分化と絶滅)が盛んに生じたことがわかります。
生物多様性の研究では、情報の不完全性(ショートフォール)が問題になります。今のところ、問題視されているショートフォールには、以下があります。
リンネアン・ショートフォール:分類情報の欠損
ダーウイニンアン・ショートフォール:系統情報の欠損
ウオーレシアン・ショートフォール:地理分布情報の欠損
プレストニアン・ショートフォール:種個体数情報の欠損
低次から高次に至るショートフォールは部分的に相関していて、そのことが高次階層におけるマクロ生態学的な生物多様性パターンの解明を阻んでいます。
また、各ショートフォールは観測が困難な部分(観測不可能領域)もあります。例えば、ミクロ進化で発端種が生まれて、マクロ進化へ至る延長種分化の過程で、分類学的に捕捉されない生物多様性(新種)が存在することもあるので、生物多様性研究にはダークマター的な要素もあるのです。
しかしながら、生物多様性ショートフォールは、最近の技術発展で、今後かなり解消されていく見込みがあります。例えば、次世代シークエンサーはリンネアン/ダーウイニンアン・ショートフォールの大部分を解消するでしょう。また、生物分布データベースや環境DNAやHabitat Suitability modelingなどは、ウオーレシアンショートフォールを劇的に解消していくでしょう。
このような技術革新によって、低次のショートフォールが解消されるに伴って、高次のマクロ生態学的パターンに関わる問題の解明が(概念的な要素も関わるので)今後より顕在化するように予想しています。
このような観点からも、深谷さんの論文は新規的な要素があります。
マクロな種プールレベルで種個体群の絶対個体数を推定したという点で、プレストニアン・ショートフォール(種個体数)の解明に成功したのですが、その過程では、大規模植生データや種の分布データを駆使して、また、新規的な統計モデルを開発して、低次のウオーレシアン・ショートフォールを克服しつつ、マクロ進化生態学的パラメータを推定しています。そして、さらに興味深い点は、高次元のプレストニアンショートフォールを解明することで(中立理論を介してではありますが)リンネアン・ショートホール解明の手がかりも与えています。
リンネアン・ショートホールはアルファ分類学の問題で、分類学者が解明すべき問題というのが一般認識ですが、リンネアン・ショートホールの実態を決定づけるパラメータ(種分化率や種寿命など)が、分類学の外のマクロ生態学的分析から示される訳ですから、興味深いです。
深谷さんの論文を元に色々考えると、生物多様性情報の不完全性問題は、より包括的なアプローチで取り組まれるべきで、ここにも進化生態学の存在意義があるように感じています。 この論文は、生物多様性科学のブレークスルーになるのではないか、と感じています。
リンネアン・ショートフォール:分類情報の欠損
ダーウイニンアン・ショートフォール:系統情報の欠損
ウオーレシアン・ショートフォール:地理分布情報の欠損
プレストニアン・ショートフォール:種個体数情報の欠損
低次から高次に至るショートフォールは部分的に相関していて、そのことが高次階層におけるマクロ生態学的な生物多様性パターンの解明を阻んでいます。
また、各ショートフォールは観測が困難な部分(観測不可能領域)もあります。例えば、ミクロ進化で発端種が生まれて、マクロ進化へ至る延長種分化の過程で、分類学的に捕捉されない生物多様性(新種)が存在することもあるので、生物多様性研究にはダークマター的な要素もあるのです。
しかしながら、生物多様性ショートフォールは、最近の技術発展で、今後かなり解消されていく見込みがあります。例えば、次世代シークエンサーはリンネアン/ダーウイニンアン・ショートフォールの大部分を解消するでしょう。また、生物分布データベースや環境DNAやHabitat Suitability modelingなどは、ウオーレシアンショートフォールを劇的に解消していくでしょう。
このような技術革新によって、低次のショートフォールが解消されるに伴って、高次のマクロ生態学的パターンに関わる問題の解明が(概念的な要素も関わるので)今後より顕在化するように予想しています。
このような観点からも、深谷さんの論文は新規的な要素があります。
マクロな種プールレベルで種個体群の絶対個体数を推定したという点で、プレストニアン・ショートフォール(種個体数)の解明に成功したのですが、その過程では、大規模植生データや種の分布データを駆使して、また、新規的な統計モデルを開発して、低次のウオーレシアン・ショートフォールを克服しつつ、マクロ進化生態学的パラメータを推定しています。そして、さらに興味深い点は、高次元のプレストニアンショートフォールを解明することで(中立理論を介してではありますが)リンネアン・ショートホール解明の手がかりも与えています。
リンネアン・ショートホールはアルファ分類学の問題で、分類学者が解明すべき問題というのが一般認識ですが、リンネアン・ショートホールの実態を決定づけるパラメータ(種分化率や種寿命など)が、分類学の外のマクロ生態学的分析から示される訳ですから、興味深いです。
深谷さんの論文を元に色々考えると、生物多様性情報の不完全性問題は、より包括的なアプローチで取り組まれるべきで、ここにも進化生態学の存在意義があるように感じています。 この論文は、生物多様性科学のブレークスルーになるのではないか、と感じています。

 RSS Feed
RSS Feed
